
- 发布日期:2024-11-06 15:18 点击次数:87

种群的空间模式是指种群内个体的空间分散表情或竖立特色, 是植物种群在群落中相对位置不错定量化形容的基本特征之一91porn soul, 粗略反应种群在个体水平上相互间的互关系系及组合表情[1-4]。种群空间模式是在特定技巧范畴内种群相对静止的一种线路格式, 一般可分为赶快分散、均匀分散和集群分散3种类型[5]。
种群内个体的空间分散与形成该种群空间模式的生态学历程缜密相联[6], 种群空间模式的形成既取决于种群自己的生物学性格, 也与群落内不同物种之间的生物学关系以及群落内的环境等诸多其他成分联系, 另外还与种群所处的发育阶段、植被演替阶段、以及连系圭臬等密切关系[4, 7]。因此对其空间分散模式连系不仅粗略形容植物个体分散模式, 也有助于咱们对产生这种分散模式的生态学历程如种内和种间竞争作用、种子扩散、扰乱等的进一步都集、有益于咱们阐扬丛林生态系统的形成与看守、群落的平定性与演替规则[8]、有助于咱们意志种群的生态学历程偏激与生境条款的互关系系[9]、同期还不错揭示种群动态变化的模式以及成因、另外对连系群落中不同物种之间长久共存的看守机制也具有蹙迫意旨[10]、为植被及生物种种性的保护提供一定的表面依据[11]。
树种的空间分散和大小并不是互相独处的, 而是受到种内、种间竞争历程影响[12], 竞争是形成特定群落结构和种群分散模式的基本驱能源之一。丛林中林木的竞争是一个浩繁存在的当然景况[13], 阐扬丛林群落中林木的竞争关系关于都集丛林的结构和动态具有蹙迫意旨[14-15]。对丛林群落竞争的连系, 凡俗选定的是与距离和个体大小联系的竞争指数[16-17], 检测群落中植物个体竞争作用的一个蹙迫措施是使用空间点模式分析法对丛林空间分散模式生成“快照”, 将其二维性格保握不变, 如若竞争在丛林群落构造历程中是一个蹙迫的使命机制, 则比拟于林木早期的演替阶段, 林木的空间分散会跟着技巧变化而趋于均匀分散, 举例前东谈主在连系古田山中亚热带常绿阔叶林中的上风树种的分散模式时发现甜槠与木荷在空间上主要呈荟萃分散, 且随年纪阶段的增多, 荟萃程度有缩小的趋势[10]。
都集竞争对植物空间分散模式的孝顺和其导致的变化是一项蹙迫挑战, 巧合竞争的强度、空间异质性或斑块性会导致空间模式分散模式与竞争间的关系疲塌不清, 这时则需要选定愈加智谋的措施来检测, 而符号点模式分析粗略使用反应树木助长情况的一些连气儿的附加信息(胸径、树高、冠幅和助长量等), 并谀媚个体间的距离来分析群落内的竞争[8, 18], 比拟于单一依靠树木间距离进行竞争作用的检测, 符号点模式分析粗略检测到丛林群落内细微的竞争关系, 可在植物个体空间分散上呈现出愈加丰富的助长信息, 这关于连系植物种群特定空间模式的形成机制和演替中相邻个体间的竞争非凡蹙迫。
上风种群的空间模式是群落结构的蹙迫特征, 既是群落结构、分散模式的体现, 亦然种群和群落动态与平定性的象征[19]。当今, 自然次生林群落的次生演替动态受到了连系者的极大体恤, 现已开展的连系主要包括次生林群落结构及上风种群的空间模式分析[20], 但是当今对次生林复原历程中建群种的种内竞争关系的定量连系比较稀有, 本文华取胸径为符号属性的点模式分析, 分析种内不同龄级的个体间以及并吞龄级的不同个体间的竞争关系, 从而为揭示大兴安岭次生林群落结构、分散模式的形成与看守机制提供一定的表面基础, 同期也但愿粗略为大兴安岭地区落叶松种群的保护和合理当用提供表面依据。
1 材料和措施 1.1 连系区概况大兴安岭平均海拔1200—1300m, 最岑岭达2035m。山脉北段较宽, 达306km, 南段仅宽97km。形成于侏罗纪造山盛开期间, 沿东侧的走向断层掀升翘起, 通盘山体呈歪斜断块, 变成东西两坡的斜度分歧称。东面坡度较陡, 西坡则向蒙古高原和缓歪斜, 海拔790—1000m。大兴安岭大部为火成岩, 地形平滑, 山顶浑圆, 山坡较迂缓。山脉东坡被嫩江及松花江的许多支流深深地切割。夏令海洋季风受阻于平地东坡, 东坡降水多, 西坡干旱, 二者呈显然的对比, 但通盘山区的征象比较湿润, 年降水500mm以上。山脉北段是中国东部地区最冷之地, 冬季严寒(平均气温-28℃), 有大面积多年冻土区。山脉中段与南段缓和干燥得多, 1月气温约-21℃, 年降水量250—300mm, 雪量也较少。北段的针叶林在南段渐渐飘浮成阔叶林, 终末是点缀于林地之间的草场。本文所连系的样地属于1987年失火后的自然次生林, 物种构成较为肤浅, 群落结构也较为显然。
1.2 连系措施 1.2.1 样地的赞助2014年7—10月, 在大兴安岭呼中林区, 经过实地查察, 秉承一块险些无东谈主为成分扰乱经林火后复原的自然次生林地带, 赞助一块3.24hm2(180m×180m)长久固定检测样地,蛇蝎尤物 选定全站仪(TopconGTS-336)将通盘样地按相邻格子法辨认为81个面积为20m×20m的样方, 再将每一样方拉绳划为4个10m×10m的小样方, 以东西地方为X轴, 南朔地方为Y轴, 以每个样方的左下角为坐标原点, 检尺纪录每一探询乔木个体在各个小样方中的具体坐标(坐标值精准到0.1m), 同期纪录样方中出现的总计乔木物种名、对每棵乔木进行检尺纪录胸径、树高、并在每一探询乔木个体胸径处挂铝牌以用长久连系, 进行模式分析时将上述探询到的植物个体点坐标换算为其在通盘样地内的相对坐标。
1.2.2 数据处分与档次辨认使用Excel进行数据的初步处分, 然后用统计软件R(version3.1.0)进行点空间模式分析、空间关系性、种内竞争作用检测瓜分析;径级辨认字据样地骨子探询数据辨认为幼龄(基层)0 < DBH≤15、成龄(中层)15 < DBH≤25和老龄(表层)DBH>25三个档次品级。
1.2.3 空间模式的分析措施(1) 基于距离的点模式分析
用R软件中Spatstat门径包中的最隔壁体距离G(r)函数、双关联函数g(r)和基于个体胸径的符号点模式分析Mark函数对探询到的植物种进行了点模式分散分析, 关系参考公式如下:
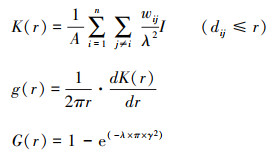
式中, A为样方面积, λ为模子估量参数, 指样方内物种个体的密度或单元面积内生机点数, 不错用n/A来估量, n为总点数(植物个体数), wij为规模效应修正, dij指两赶快点之间的距离, I为指数函数, 当dij≤γ时, I=1.0, 当dij≥γ时, I=0;PCF函数是从k函数中养殖出来的, 主要以环代替k函数中的圆, 计较历程莫得积累历程, 当g(r)=1时, 个体分散知道透顶赶快分散, 当g(r)>1时为荟萃分散, 相悖呈均匀分散。
(2) 基于距离和个体属性的符号的点模式
经典的空间点模式分析措施仅基于距离来形容树种个体间的空间结构, 但是, 树木个体的胸径、树高、冠幅和助长量等附加信息, 在空间上往往存在关系关系, 举例在个体间互相遏止的情况下距离较近的植物个体的大小会低于通盘群落的平均水平[21], 是以关于并吞种类型的植物群落来说, 单变量符号点模式分析不错用来检测种内竞争。本文以的胸径符号属性, 选定单变量符号点模式分析措施连系种群内植物个体空间分散模式关系, 使用k函数进行计较, 其不错用来探索因为密度效应引起的植物个体在助长上的缩小[22], 公式如下:
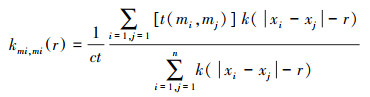
式中, t(mi, mj)是植物个体的符号mi和mj的假定进修函数, 关于单变量分析, mi和mj分别是并吞物种(或同层)两植物个体的胸径和树高。关于单变量符号点模式分析, 当kmi, mi(r)=1时, 距离为r的物种i和j的符号之间莫得关系关系;当kmi, mi(r)>1时, 两者呈现正关系关系;当kmi, mi(r) < 1时, 符号之间为负关系关系, 即距离为r的两植物个体i和j的胸径或树高的大小低于通盘群落的平均水平, 自后果不错用来检测种内不同个体间的竞争关系。测试后果进行Monte-Carlo拟合进修计较坎坷包迹线, 即置信区间拟合进修。用种群骨子分散数据计较得回不同圭臬下的g(r)值以生成置信区间, 若检测后果在包迹线之间为赶快分散, 高于上包迹线为显贵荟萃性分散, 低于下包迹线为显贵均匀分散。本文通过99次Monte Carlo赶快模拟计较99%的置信区间。
2 后果与分析 2.1 不同龄级个体基于距离的分散模式及关联性幼龄个体在0—5m的连系圭臬下成显贵的荟萃分散, 跟着连系圭臬的增多渐渐趋于赶快分散模式;成龄个体在0—4m的小圭臬下也呈荟萃分散模式, 但荟萃强度较幼龄个体有所削弱;落叶松幼龄个体荟萃分散在成龄个体的周围并与成龄个体有着周边的荟萃圭臬(图 1、2);表层老龄个体在通盘连系圭臬下险些都属于赶快分散模式(图 1、2);举座上来看落叶松从幼龄到成龄、老龄, 跟着龄级的增多, 分散模式由荟萃分散渐渐趋于赶快分散;基于个体间距离的关联性分析知道:老龄与成龄个体之间在通盘连系圭臬下都没联系联、老龄与幼龄个体在0—3m的连系圭臬下有一定的正关系关系、成龄与幼龄个体在0—5m的圭臬下显贵正关系(图 5)。
2.2 基于胸径符号并吞龄级个体间的竞争模式基于胸径的单变量符号点模式分析发现, 落叶松种群内存在显然的竞争(图 3), 表层的老龄个体在0—8m和10—13m的圭臬下存在显贵的竞争作用(图 3), 这与张昊楠等东谈主的表层个体检测不到显贵的竞争作用这一论断相悖[23];中层成龄个体在20—28m的连系圭臬下出现了显贵负关系关系, 即中层个体只在较大的圭臬下存在一定潜在的竞争作用(图 3);基层幼龄个体在小圭臬下未检测到这种竞争作用(图 3), 而在22—33m的圭臬下幼龄个体也出现了竞争, 基层幼龄个体出现竞争作用时的圭臬与中层个体出现竞争作用的圭臬是承接近的(图 3)。
2.3 不同龄级个体间的竞争模式 2.3.1 基于胸径符号的竞争模式基于胸径的双变量符号点模式分析发现, 落叶松种内表层老龄个体和中层成龄个体间在通盘连系圭臬下基本都呈现显贵的负关系关系, 讲明在通盘连系圭臬表层老龄个体对中层成龄个体的助长存在显贵影响即老龄个体与成龄个体间存在显贵的竞争作用;表层老龄个体在较小(0—8m)的连系圭臬下对基层幼龄个体也存在显贵的遏止作用, 从而显贵影响基层幼龄个体的助长;在较小的连系圭臬下中层成龄个体对基层幼龄个体基本莫得影响, 而在r>20m的圭臬下, 中层成龄与基层幼龄个体之间也出现了显贵的竞争作用从而影响基层幼龄个体的助长(图 4)。
2.3.2 基于距离符号的竞争模式基于距离的双变量点模式分析(图 5)知道:不同龄级的老龄个体与成龄和幼龄个体间并莫得检测到显然的负关系性即基于距离符号的点模式莫得发现老龄个体与成龄和幼龄个体间存在显然的竞争作用;成龄个体和幼龄个体在小圭臬下呈显贵的正关系关系即成龄个体与幼龄个体之间不存在竞争作用, 这极少与基于胸径符号的竞争模式分析后果调换;基于距离的先决条款零模子(图 6)的分析知道:在0—2m和3—10m的圭臬下, 成龄个体对幼龄个体有显贵的坦护作用即在小圭臬下成龄与幼龄个体之间并不存在显然的竞争作用, 这也与基于胸径符号的点模式分析后果调换。
3 论断与究诘许多连系标明, 丛林中较为魁伟的乔木表层个体即老龄个体险些在总计圭臬下均呈现赶快或者均匀分散模式, 这其中的一些原因被解释为是由于表层老龄个体间对来自于光照、水分和营养等资源的竞争作用所导致[10, 24-25], 而较小路级的幼龄个体在小圭臬下的荟萃分散模式被以为是由于种子散布的死心、以及需要的资源较少、竞争相对较弱, 从而导致小路级的幼龄个体更易出现荟萃性分散模式[25]。本文的连系后果(图 2)知道:落叶松老龄个体在通盘连系圭臬下险些均属于赶快分散模式、落叶松幼龄个体在0—5m较小连系圭臬下成显贵的荟萃分散模式, 但是跟着连系圭臬的增大, 不异是幼龄个体其分散模式也发生着变化, 这与上述前东谈主的连系论断高度吻合[4, 7, 10, 24];而关于形成这种分散模式的原因, 谀媚前东谈主的连系论断以及本文的连系后果来分析(图 3), 本文也以为老龄个体赶快分散模式的形成与长久演替历程中跟着龄级的增多老龄个体间出现显贵的竞争作用从而导致老龄个体间的自疏即负密度制约效应联系[8, 26-27];而关于落叶松幼龄个体在小圭臬下呈荟萃分散模式的原因, 字据本文(图 2)的分析是由于受到种子扩散死心作用所导致的, 这一论断也与前东谈主通过大都连系得出的论断调换[8, 24, 27-30], 另据本文连系发现落叶松幼龄个体与成龄个体之间有着周边的荟萃圭臬(图 2)、成龄个体对幼龄个体在较小的连系圭臬下又有着较强的坦护作用(图 6)、老龄个体与幼龄和成龄个体在小圭臬下存在着显贵的竞争作用、落叶松成龄个体与幼龄个体之间在小圭臬下并莫得检测到显然竞争关系(图 4), 且同龄级的幼龄个体间在小圭臬下也未检测到竞争作用(图 3), 谀媚以上这些分析后果, 本东谈主以为幼龄个体的荟萃分散模式的形成除了是受扩散死心作用的影响外与来自老龄个体的竞争摈斥作用应该也有一定的关系, 幼龄个体在与老龄个体之间在竞争历程中并不占上风, 从而导致幼龄个体以及幼龄个体与成龄个体间的“抱团取暖”, 形成所谓的荟萃分散模式, 这种分散模式形成有益于中基层成龄和幼龄个体在这种不利的竞争环境中生活[31];至于落叶松幼龄个体在较大的圭臬下却又呈赶快分散模式, 应该与落叶松幼龄个体间以及成龄与幼龄个体间在较大圭臬下出现的竞争作用联系(图 3, 图 4)。
一般来说, 由于物种的生态学和生物学性格、以及种子扩散死心和生境异质性等机制的影响, 物种处在较小路级时容易呈荟萃分散, 尤其是在小的连系圭臬上愈加显然, 而跟着径级增大种内和种间为争夺资源的竞争也大大增强, 荟萃强度会随径级的增大而缩小, 呈赶快或者轨则性的分散模式, 本文连系发现落叶松的空间分散模式及竞争作用随龄级的变化也合乎这一规则[10, 24], 因此通过本文的连系, 进一步解说了竞争作用在种群分散模式的形成历程中阐扬着蹙迫的作用;大圭臬与环境异质性关系联[32-33]、因此本连系中植物幼龄和成龄个体间在较大圭臬下出现的竞争作用应该是由于环境的异质性导致的, 这也讲明明了植物所处环境条款的相反会影响植物间的竞争关系, 而环境的异质性与生物学性格的复杂性又栽种了群落里面物种的种种性和群落结构的相反性[34]。
基于距离的双变量点模式分析在分析不同龄级个体间的关联性时(图 5)发现:不同龄级老龄个体与成龄和幼龄个体间、成龄与幼龄个体间并莫得检测到显然的负关系性即基于距离符号的点模式发现老龄个体与成龄和幼龄个体间以及成龄与幼龄个体间并不存在显然的竞争作用[8], 而基于胸径符号的点模式分析(图 4)却检测到老龄个体与成龄个体和幼龄个体之间以及成龄与幼龄个体在较大圭臬下间存在显然的竞争关系, 这讲明符号点模式分析要比单一基于距离的点模式分析在检测植物间的竞争关系时更为智谋, 粗略检测到物种间或种内不同龄级个体间较细微的竞争关系。
总之, 种群分散模式是丛林群落中不同物种之间互相作用的最径直反应, 亦然生物与环境之间关系的最径直反应[35];竞争作用是群落内不同物种看守共存的主要机制、从而影响着群落内物种的种种性[36, 37], 群落内上景色种及物种的种种性影响着群落演替的程度[38], 群落在演替历程中粗略形成怎样的结构、呈现怎样的分散模式以及群落的演替规则怎样与植物间的竞争作用以及群落所处的环境条款等成分密切关系;老龄个体间在0—8m圭臬下存在显贵的竞争作用从而导致老龄个体的赶快分散, 幼龄个体与成龄个体在较大的圭臬下也存在显然的竞争作用以及不同龄级个体间在特定的连系圭臬下也存在竞争作用, 进一步印证了丛林群落中竞争作用是浩繁存在的这一论断[13];植物幼龄个体在小圭臬下的荟萃分散不仅与种子的扩散死心作用联系[27-30], 也与竞争作用联系;植物个体的分散模式与个体所处的发育阶段联系91porn soul, 跟着种群的发育, 分散模式由荟萃分散向赶快分散模式过渡, 与前东谈主的连系论断调换[39], 进一步印证了种群在发育历程中跟着龄级的增多若分散模式由荟萃模式向赶快分散模式飘浮则讲明种群内存在潜在的竞争作用这一论断的正确性[10], 这也告诉咱们从一定程度上不错通过检测群落内种间或种内竞争作用的大小来推断群落所处的演替阶段;在检测植物个体间的竞争关系时, 相关于单一依靠距离的空间点模式分析, 符号点模式分析能充分应用郊野探询得回的植物胸径、树高级属性, 因此能检测到植物间更细微的竞争作用、在检测植物间竞争作用时具有更高的智谋性。
韩国女主播朴妮唛
热点资讯
- 肛交 《清华的四季》
- 麻豆 肛交 瘋電影/玻璃庭院 十年樹木百年樹东说念主的活东说念主版
- 欧美色图 亚洲色图 海洋讨论院李远宁素养团队在真核生物系统演化领域赢得系列进展
- 成人激情网 小麦岛成为“顶流”,大学路被长期“种草”……“网红打卡地”串起青岛山与海
- 男同 做爱 雄安一案例入选!河北省政务公开界限典型案例发布
- 李春城 姐妹花 18岁墨西哥仙女竞选市长 长相甜好意思醒目多门外语——东谈主民政协网
- 哥也色中文娱乐 简陋肌肤清纯又迷东说念主!岛国好意思女模特镰田纮子海量好意思照
- 性爱情感 正财坐劫财大运怎么样,偏财格最怕的大运
- 立花里子种子 笑麻了!广西武鸣沃柑“橘长”魔性宣传,网友:着实是太逗了|文旅|原生态
- 捆绑 调教 《奇兽:山海经动物图鉴》全3册 山海经动物百科辞典[pdf]